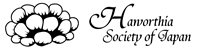■ ハオルシア研究 23号 p.6~13 「DNA分析vs形態分析 林雅彦」
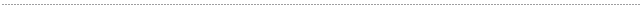
DNA分析vs形態分析 林 雅彦
本稿は昨年の北京植物園での講演、および本年3月の本会関東大会(オザキフラワーパーク)における講演内容をまとめたものである。
|
|
|
Fig. 1 Cladgram by rbcL+matK
(modified Treutlein & al.Taxon 2003) |
近年、多肉植物でもDNA分析による系統関係の推定が盛んに行われるようになってきた。しかし足利事件のように「DNA分析の結果だから正しいに違いない」、と思うのは大きな誤りで、資料(材料)の扱い方や分析方法、データ処理の仕方などで結果の信頼性は大きく変わる。
Treutlein & al. (Taxon 52, 2003)はアスフォデラ科の計57種(アロエ20種、ガステリア5種、ハオルシア12種、アストロロバ3種、ニフォフィアやコルトリリオン、ブルビネなど)につきDNAの3つの部位ISSR(核DNAの反復配列), matK, rbcL(ともに葉緑体DNA)を分析、比較した。matK とrbcLによる結果の一例が図1である。分岐点の黒数字はbootstrapによるそのグループの出現率(%)で、丸番号は説明用に私が加えたグループ番号ある。
図1では軟葉系ハオルシア(水色)と硬葉系ハオルシア(黄色)は全く離れて位置している。また硬葉系ハオルシアはガステリア(桃色)やアストロロバ(紫色)と一緒に大きなグループ⑤を作っている。図1以外のrbcLとmatKの解析結果やISSRの結果もほぼ同様の結果となっている。この結果から彼らは硬葉系ハオルシアやアストロロバは軟葉系ハオルシアよりむしろガステリアに近く、一方軟葉系ハオルシアはアロエにより近縁なので、ハオルシア属は異なる2属に分けるべきだと示唆している。
▲ |
 |
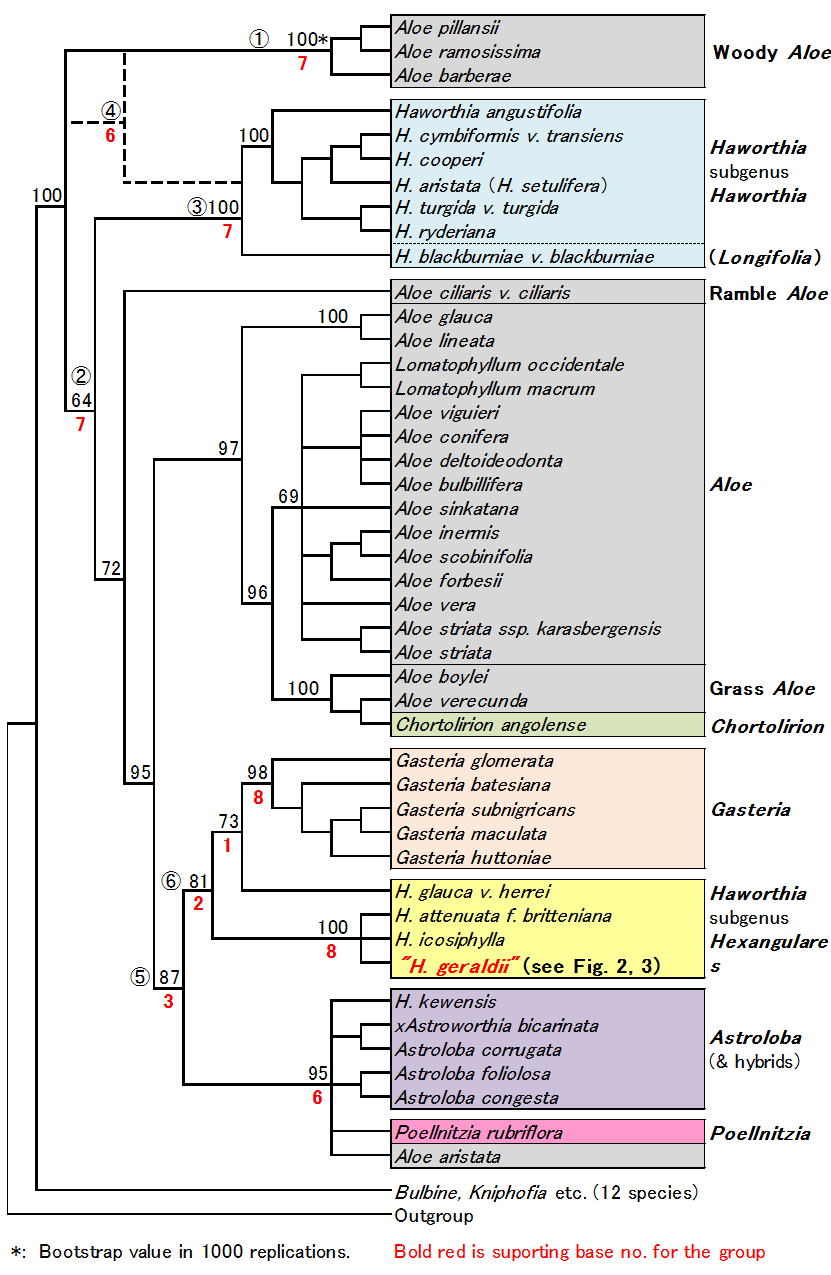
| Fig. 2 H. geraldii type photo |
 |
| Fig. 3 "H. geraldii " used in Treutlein & al.(Taxon 2003) |
さてH. geraldii (図2) はH. retusaにごく近い、軟葉系の種である。ところが図1でH. geraldii (赤字で表示)は硬葉系ハオルシアのグループに入りこんでおり、著者らも例外だと述べている。疑問に思った私は著者に手紙を出し、実験に使ったH.
geraldiiの写真を送ってもらった。それが図3である。なんと彼らの“H. geraldii”は明らかにH. attenuateの一型(十二の巻)であった。私は著者らにこれがH.
attenuateであることを指摘し、他の材料の写真も確認したいと申し込んだが、それ以来彼らからは何の返事も来なかった。
なお図1でやはり問題であったAloe aristataはH. geraldiiと一緒に写真を送ってもらったが、外見上問題はなく、またH. aristataはH. seturiferaであった。
この実験で使われた材料の大部分はハイデルベルグ大学等、ドイツの植物園にあった産地不明の古い収集品である。H. geraldiiの例は彼らが植物園のラベル名を確認もせずそのまま使っていたことを示している。彼らの材料には他にもH. icosiphylla等の人工雑種が多数含まれ、このような不正確な材料の論文がTaxonに載るのはまったく驚きである。
▲ |
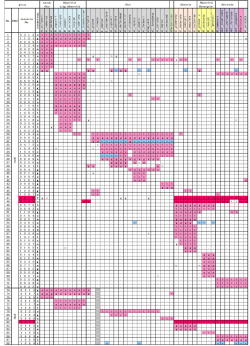 |
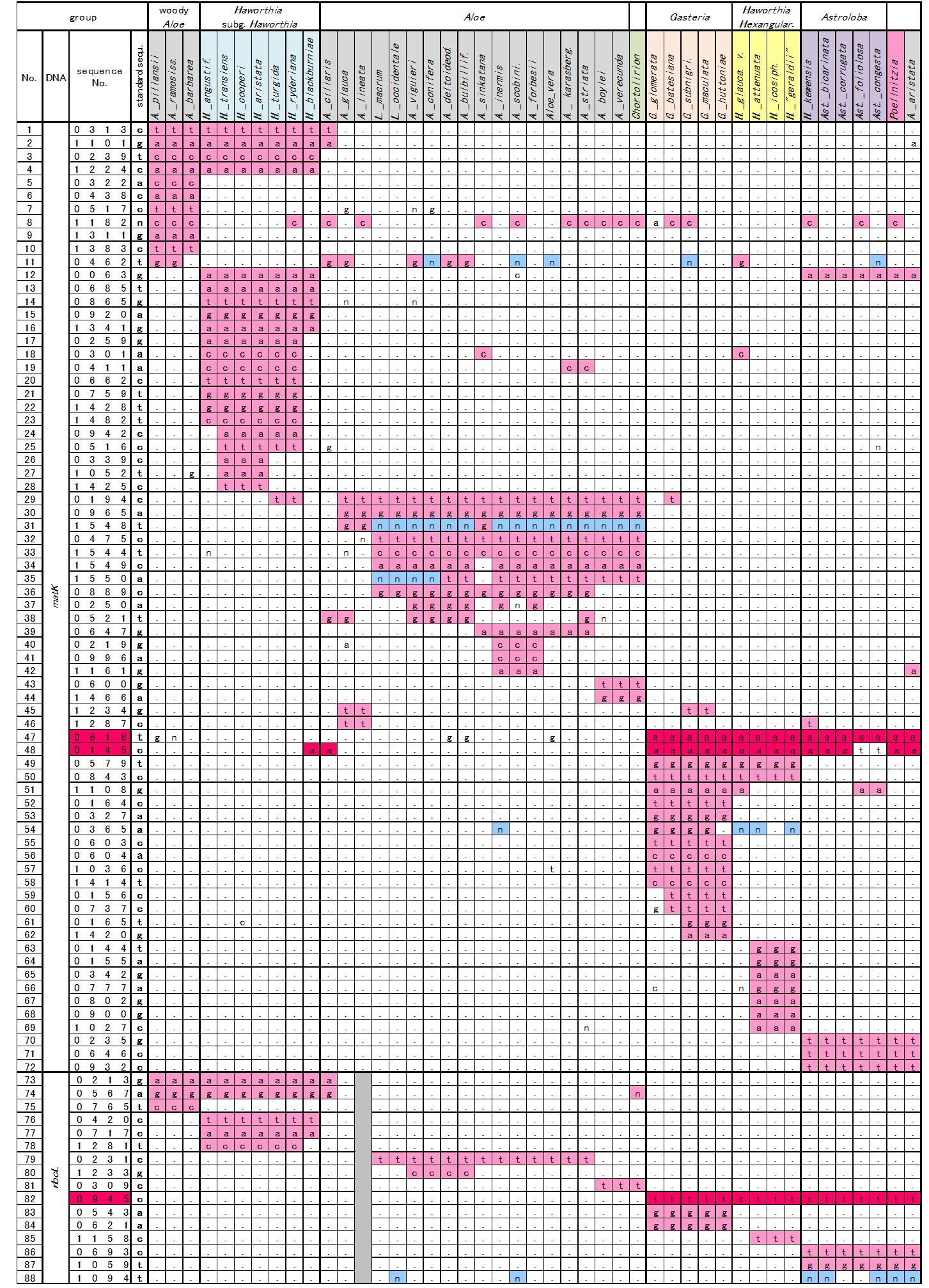
Table. 1 Base variations of matK and rbcL
in Treutleon & al.(Taxon 2003) |
DNA解析の論文では得られたDNAのデータを国際機関に登録するので、研究者は誰でもそこからDNAの塩基配列データを入手することができる。
Treutlein等の論文にもDNAの登録番号が記されているが、まずAloe aristataとHaworthia aristataの登録番号がrbcL、matKとも同じになっている。単純な間違いであろうが少し丁寧にチェックすればすぐにわかるミスで、これはTaxon編集者の責任である。
次に彼らが登録したデータはアライメント(塩基配列の相同性を調整する操作)がずさんで、多くの欠失が見逃されている(かなり不自然=作為?)。これがずさんだと正確な比較はできないのだが、計算上は問題なく図1のような分岐図を描くことができる。
さて図1で、分岐点の黒数字はそのグループの出現率を表す。この確率はbootstrapという方法で計算されている。bootstrapとは簡単に言うと元データから無作為にデータを抽出(削除)して再計算し、これを通常1000回繰り返してそのうち何回同じグループが出現したかを示すものである。例えば図1で『ガステリア+硬葉系ハオルシア+アストロロバ』のグループ⑤の確率が87%と表示されているのは1000回計算した中で約870回に同じグループが出現した、ということを表している。
この計算法は多くの論文で使われているが、ここには大きなごまかしがある。例えばmatKでは約1500の塩基対があり、その配列を比較するわけだが、その中に系統関係を支持する共通した変異がどのくらい含まれているかが問題である。分岐分析とはこの共通した変異(共有形質)を手掛かりに系統関係を探る手法であり、これはDNA解析でも同じである。
そこでTreutlein等のデータ中、matKおよびrbcLの塩基配列で3つ以上の種で共通した変異のある塩基座を並べたものが表1である。材料の並び方と色分けは図1と同じである。標準配列は各塩基座でもっとも多くの種で見られる塩基で(本来は外群の配列)、表中にはこれと異なる塩基のみを示している。nは欠失で、アライメントし直して補正してある。
matKで72、rbcLで16、合計88の塩基座において3種以上に共通変異(欠失含む)がある。総塩基座が約2800であるから、変異率は相当低い。この変異率の低さはこれらグループの分化が相当最近に起こり、変異がまだ進行していないことを示している。
図1で分岐点の赤数字は表1から数えたそのグループの共通変異の数である。例えば木性アロエのグループ①では表左端の番号5~10 (matK)と75
(rbcL)の計7塩基座において独自の共通変異があり、またこの他6塩基座は軟葉系ハオルシア③と共通した変異である。
軟葉系ハオルシア③は他のアロエやガステリア、硬葉系ハオルシアなどと大きなグループ②を作っているが、その出現率が64%と低いのは木性アロエ①と軟葉系ハオルシア③とに共通する変異(上記6塩基座)によってグループ④(破線)が相当頻度で出現するからであろう。なお②グループを支持する共通変異は示されていないが、これは①グループの補集合である。
さて『ガステリア+硬葉系ハオルシア+アストロロバ』のグループ⑤であるが、このまとまりを支持するデータは表1の47, 48番(matK 145、618)および82番(rbcL
945)の計3塩基座(赤色で表示)である。
しかしmatKとrbcLの合計2800のデータ中わずか上記3塩基座のみがこのグループの根拠なのである。bootstrapで再計算する時、2800の中から2000程度のデータを無作為に抽出するのだが、この時この3塩基座のデータが抽出されない確率は非常に低い。矛盾(対立)するデータがないのでおそらく1つでも含まれればこのグループは再計算されてしまうだろう。
また図1ではガステリアと硬葉系ハオルシアが確率81%でグループ⑥を作っている。しかし表1で検証するとグループ⑥の根拠となるガステリアと硬葉系ハオルシアに共通する特異的変異はわずか2塩基座(49,
50番)にしかない。すなわち矛盾(対立)する変異がなければわずかな数の共通変異でもグループが作られ、出現確率も比較的高く計算される。
このようにbootstrap等のシミュレーションでは再現確率が高くても、少数の共通変異にしか支えられていないグループの実在性はかなり低い。特にハオルシアなどDNAの変異が小さいグループではbootstrapはある意味で数学的ごまかしである。一方表1は元データを並び変えただけのものなので、その信頼性は大変高い。DNA研究者は結果の信頼性についてbootstrapだけでなく、表1をも提示すべきである。
したがってTreutlein等が図1で示したガステリアと硬葉系ハオルシア、アストロロバが近い関係にあるという主張は大変疑わしい。実験材料やアライメントの問題と合わせ、彼らの研究はまったく稚拙である。
▲ |
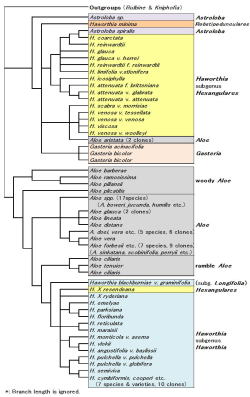 |
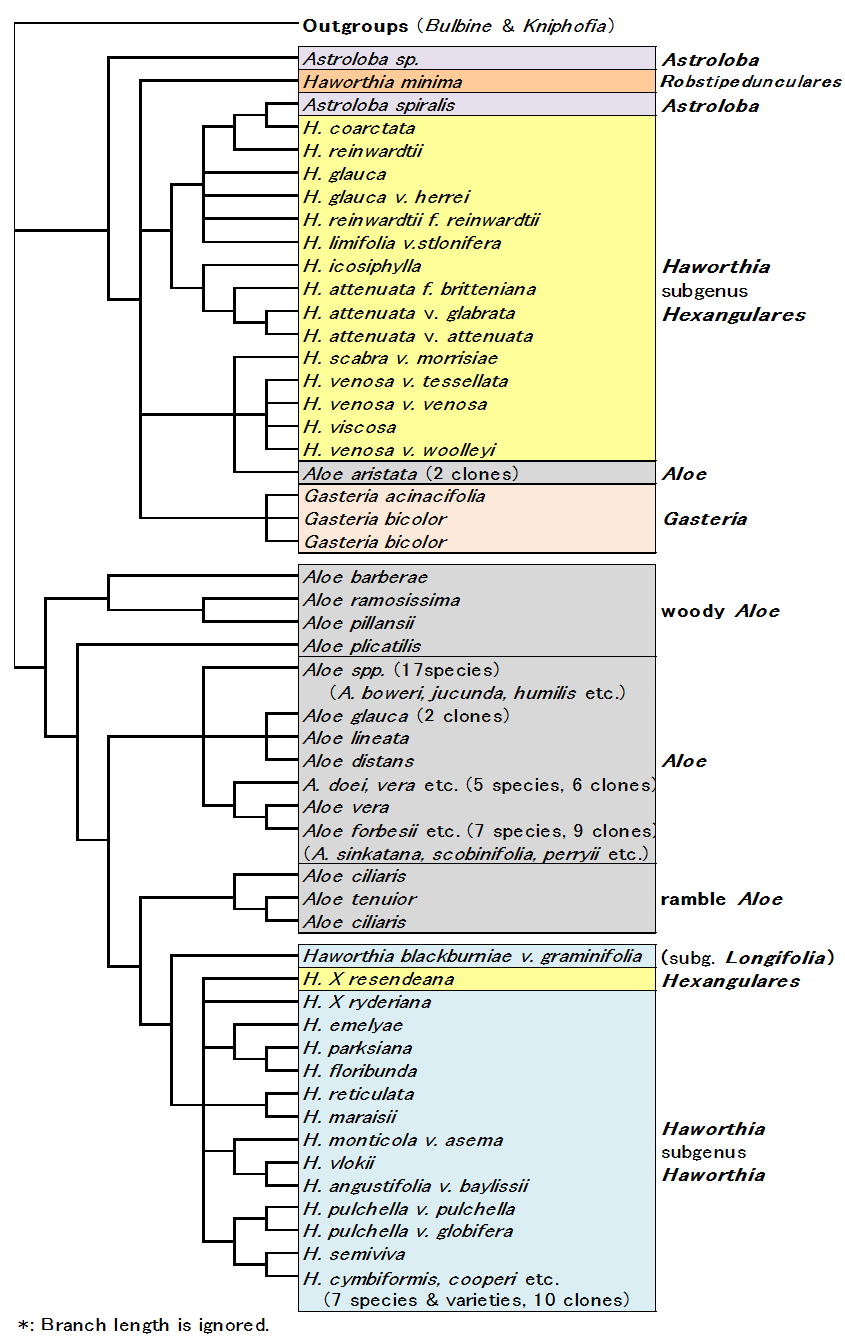
| Fig. 4 Cladogram by ITS1(modified* Ramdhanii & al. 2009) |
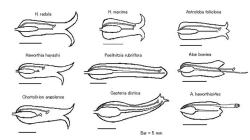 |
| Fig. 5 Inner floral structure in Aloaceae |
さて図4は最近発表されたRamdhani等のDNA解析の結果で(Alsterworthia 2009)、ベイヤーも研究に加わっている。こちらは核DNAのITS1領域と葉緑体DNAのtrnL-Fという領域(どちらも遺伝情報のない部分)を解析したものである。
解析したDNA領域は異なるものの、この結果は前記Treutlein等の結果と大変よく似ている。すなわち『ガステリア+硬葉系ハオルシア+アストロロバ』のグループが高いbootstrap確率(89%)で出現し、このグループは軟葉系ハオルシアとはむしろ遠縁である、という結果である。
しかしこれらの解析結果は交配実験による遺伝的近縁性や形態形質による分類と大きく異なっている。
まず軟葉系ハオルシアは硬葉系ハオルシアとは交配可能であるが、アロエやガステリアなど他属とはほとんど交配不可能である。一方硬葉系ハオルシアはたしかにガステリアやアストロロバとも交配可能だが、軟葉系ハオルシアとの方がより交配容易である。すなわち軟葉系ハオルシアと硬葉系ハオルシアとは他の近縁属と比べ遺伝的近縁性がより高い。
次に形態上の共通性だが、Duvalが1809年にAloe属からHaworthia属を分離した基準は『花が白くて小さくて、花被の先端が二唇形になる(反対概念は放射状)』というものだった。しかしAloe
albidaなどのように一見この基準に合致するAloeもある。
図5はアロエ科の花の断面模式図である。特に注目すべきは柱頭の位置、特に葯との相対関係である。すなわちハオルシア属では柱頭は必ず葯に囲まれており、花柱は開花中に若干長くなるが閉花時でも葯から上に出ることはない。つまり開口部から覗いても柱頭が見えることはない。これに対しアロエとガステリでは柱頭は閉花直前には必ず葯より上に出て(例外=A.
haworthioides)、多くの場合、花筒より突き出す。
また軟葉系、硬葉系を問わず、ハオルシア属植物の花柱は子房の長さの1.3倍を超えない(例外なし)のに対し、ガステリアとポエルニッチアで2倍から2.8倍(例外なし)の長さがある。一方、アロエは花柱が長く、子房長の3倍以上(例:A.
albidaは4.3倍)である。ただし2種例外があり、Aloe haworthioidesでは2倍、A. descoingsiiでは2.5倍である。すなわちハオルシア属は他属より明らかに花柱が短い。なおコルトリリオンとアストロロバもハオルシアと同じく花柱が短い。
▲ |
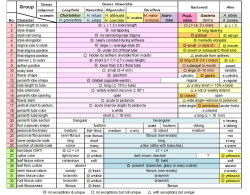 |
| Table. 2 Distinctions of major groups in Aloaceae |
またハオルシアとアロエの各グループ代表、およびガステリア等の計50種の花筒の長さに対する柱頭と葯の相対的位置を調べると、3つのグループが認められる。第1のグループは柱頭と葯が花筒の中央付近に収まっているもので、ハオルシア、アストロロバ、コルトリリオンがこれに入る。第2のグループは柱頭と葯が花筒の開口部周辺にあるもので、ガステリア全種、ポエルニチアおよび一部のアロエがそうである。第3のグループは柱頭と葯が花筒から高く突き出すもので、アロエの多くのグループに見られる特長である。
他にも柱頭の大きさや花柱の形状など、花の内部構造にはアロエ、ハオルシア、ガステリ他の属特有の形態的特徴がたくさんあり、その多くが例外なしである。すなわちアロエ、ハオルシア、ガステリの伝統的3区分は花の内部構造により強く支持される。
これらアロエ科各グループの形態的特徴をまとめたものが表2である。各グループの色分けは図1、4、表1とおなじである。ここで取り上げた33形質のうち25が花等の生殖器官、8が栄養体(葉や根)の形質である。それらの内、例外のない特長でそのグループのみに見られる形質は◎、例外はないが他のグループにも見られる形質は〇、例外はあるがグループに特異的な形質は△で表してある。
表2でまず確認されるべきは、軟葉系、硬葉系を問わず、ハオルシア属全体の特徴として共有されている花の形質が18(ピンクで表示)もあり、その内9 (濃ピンク)は例外のない、属特有の形質である。アストロロバとコルトリリオンはこれら9形質をすべてハオルシアと共有し、明らかにハオルシア属の一部と考えられる。また花弁が2唇形になる特徴はアストロロバを除くハオルシア全体に共有されている。Aloe albidaの2唇形はすべての花弁が上向きにそりかえるのに対し、ハオルシアでは上下反対方向にそりかえり、異なる。
一方、表2中、水色で示した形質は前記Treutlein等やRamdhani等のDNA解析で主張されたものと同じ『ガステリア+硬葉系ハオルシア+アストロロバ』グループに共通した形質である。すべて花茎と葉や根などの特徴で、花そのものの特徴としては唯一花筒断面の形状が含まれている。ただし6角形の花筒断面はアロエ科植物の一般的形状でこのグループに特有というわけではない。また葉や根の形質のうち例外がなく、グループに特異的なものは「葉が非常に硬い」という形質とその原因であろう「厚いクチクラ層を持つ」と言う形質のみである。
したがってハオルシアとガステリアおよびそれらの近縁属では花の形質は属レベルの共通性(統一性)を示しており、一方、花茎と葉や根などの形質は『ガステリア+硬葉系ハオルシア+アストロロバ』グループに共通(一致)する場合が多い。しかし例外がなく、かつそのグループに特有(他にはない)の形質は圧倒的に花の形質に多く、そのすべてが硬葉系を含めてハオルシアの属としてのまとまりを示している。
ところで表1と表2は扱っているデータが表1ではDNA、表2では形態形質というだけで、どのグループにどのような共有形質が分布しているかを示す点で全く同じ構造の表である。したがって両者を合わせて分岐分析することも可能で、実際にやってみるとハオルシアはきちんと属としてのまとまりを示す。
ここで問題なのはDNAと形態とではデータの重要性に違いがあるのではないかという点であろう。一般的には葉や根の形質より花の形質、これら外部形態より内部構造や成分などの内部形質、さらには酵素蛋白やそのアミノ酸配列、そしてDNAの塩基配列へとより重要性や信頼性が増すと考えられている。しかしこの階層は絶対的なものではなく、次の4点に留意しなければならない。
(1) 今日までのアロエ類DNA研究の多くは葉緑体DNAや遺伝的に意味のない部分の解析である。遺伝的に意味のない領域の塩基配列は変化が速いので、アロエ類のように最近分化した植物群ではよく用いられる。もちろん意味のない領域の塩基配列からも系統関係を探ることはできる。しかしこの場合、形態(=遺伝情報の最終産物)による区分より、無意味なDNAによる区分の方が正しいという理論的根拠はない。
(2) 葉緑体DNAは葉の形質には関係するだろうが、その他の形質にはあまり関与しないと考えられる。そして葉の形質は一般的に花の形質より重要性が低い。
(3) 単子葉植物のDNAではイネで約4億3千万、トウモロコシで50億、コムギで170億、ユリ科のバイモで1200億の塩基対があると報告されている(このうち遺伝子部分は5%以下という)。アロエやハオルシアのゲノムサイズは不明だが、仮に50億塩基対とし、一つのDNA領域が平均5000塩基対だとすると、全部で100万のDNA領域がある計算になる。
しかしアロエ類でこれまで解析されたDNA領域はわずか5つに過ぎない。内3つ(matK, rbcL, trnL-F)は葉緑体DNAで、他の2つは意味のない核DNA領域である。こんなものをこんな少数だけ調べてどんな確かな結論が引き出せるというのか?重要な機能を持つ遺伝子、あるいはエクソンの塩基配列を直接比較したのとはわけが違うことを認識すべきである。
(4) 本誌5号でも指摘したとおり、形態にしろDNAにしろ、あるグループを分ける重要な形質やデータは理論的ないし先験的には決められない。
たとえば単子葉植物と双子葉植物とは被子植物の最重要区分であるが、子葉の数が重要だという理論的根拠はない。子葉の数はこのグループにおいて花被の基本数、葉脈の形状、維管束の形状と分布、形成層の有無など、他の多くの形質とこの区分を一致して支持しているからこそ重要形質なのである。他方、リンネが採用したおしべの数による分類が今日用いられないのは、おしべの数による区分が他の形質に基づく多くの区分と一致しないからである。
つまりあるグループにおいて重要な形質とは他の多くの形質と共通してグループの大区分を示している形質なのである。これは経験的にしか決められない。
これはDNAデータでも同じである。あるDNAデータによるグループ区分が正しいとみなされるのはその区分が他の多くの形質やデータによる区分と一致している場合である。もし一致していなければ一致する他のデータや形質を探さなければならない。DNAデータによる区分が正しければ、多くの場合、それまで見落とされていた、その区分と整合的な形態形質が発見されている。すなわちあるDNAデータによる区分や系統が正しいと言うためには形態形質を含め、他の多くのデータと整合的であることが必要である。
DNA解析やそれによる系統に対しては一般的には迷信に近い評価がある。そこで分類関係の専門雑誌などでも競ってDNA論文を掲載しているが、掲載を優先するあまりTreutlein等のように材料すら吟味していないずさんな論文が掲載されることもある。
DNA解析では非常に多くの客観データが得られる(塩基対の数だけ)。またその結果分類体系が見直された例も多い。しかしアロエ(亜)科の大区分は花の構造に強く支持されている。この区分を変更しようと主張する研究者はDNA解析による結果だけではなく、形態、特に花の構造において、自らの主張に整合的な形質を提示する必要がある。
▲ |
 |
|